
Science:揭示海马体神经回路网络形成中的轴突吸引和排斥机制
来源:本站原创 2021-06-20 06:57
2021年6月20日讯/生物谷BIOON/---海马体(Hippocampus),又名海马回、海马区、大脑海马,位于大脑丘脑和内侧颞叶之间,主要负责长时记忆的存储转换和定向等功能。海马体是被称作“海马区”(hippocampal region)的大脑边缘系统的一部分。海马区可分为齿状回(dentate gyrus)、海马体、下托(subiculum)、前下托
2021年6月20日讯/生物谷BIOON/---海马体(Hippocampus),又名海马回、海马区、大脑海马,位于大脑丘脑和内侧颞叶之间,主要负责长时记忆的存储转换和定向等功能。海马体是被称作“海马区”(hippocampal region)的大脑边缘系统的一部分。海马区可分为齿状回(dentate gyrus)、海马体、下托(subiculum)、前下托(presubiculum)、傍下托(parasubiculum)、内嗅皮质(entorhinal cortex)。这之中齿状回、海马体、下托的细胞层为单层,合称“海马结构(hippocampal formation)”,其上下夹有低细胞密度层和无细胞层。
我们感知和驾驭世界的能力需要大脑中神经回路的精确组装和功能。在发育过程中,称为轴突的神经元投射在分子线索的引导下,从大脑的非目标区域向它的目标区域延伸,在那里轴突与神经元搭档建立突触连接。在过去的几十年里,科学家们已经鉴定出几种候选的分子线索;然而,对于不同的细胞表面分子集合是否介导了对目标区域的吸引和对非目标区域的回避,仍然存在疑问。在一项新的研究中,来自美国斯坦福大学和哈佛医学院的研究人员在小鼠中发现,轴突吸引和排斥在海马体的神经回路组装过程中由同一个细胞表面分子引导。相关研究结果近期发表在Science期刊上,论文标题为“Reciprocal repulsions instruct the precise assembly of parallel hippocampal networks”。论文第一作者为斯坦福大学的Daniel T. Pederick博士。

海马体含有CA1、CA2和CA3亚区,CA1亚区中的神经元投射到下托中的目标区域。CA1到下托(subiculum)的神经元投射是沿着内侧到外侧的解剖轴组装的。这样,在这种神经回路网络的内侧部分,CA1近端(位于与CA2区域的边界附近)的神经元投射到远端下托(离CA1边界最远的部分),而在这种神经回路网络的外侧部分,CA1远端神经元投射到近端下托。
Pederick及其同事们之前的一项研究已表明,一种叫做teeneurin-3(Ten3)的细胞表面分子既被近端CA1神经元表达,也被其目标区域(即远端下托)的神经元表达;它显示,Ten3分子相互粘连,这种结合导致表达这种蛋白质的神经元之间的吸引。通过这种相互作用,表达Ten3的神经元投射被吸引到表达Ten3的目标区域。Pederick等人假设,一种类似的机制--蛋白质结合导致表达它的神经元投射被吸引到表达这种相同蛋白质的目标区域--可能参与指导外侧海马体神经回路网络的形成。
在这项新的研究中,Pederick等人使用一种叫做单细胞RNA测序的技术来分析发育中的小鼠海马体中单个细胞的基因表达,发现细胞表面蛋白latrophilin 2(Lphn2)在远端CA1神经元投射和外侧海马体神经回路网络的近端下托目标神经元中都有表达。他们最初研究了Lphn2-Lphn2的粘附和吸引是否会以类似于Ten3-Ten3粘附和吸引的方式,指导海马体神经回路的形成。然而,情况并非如此:当这些作者在非粘附性的细胞系中过度表达Lphn2时,这些细胞并没有相互粘附在一起。相反,表达Lphn2的细胞很容易与表达Ten3的细胞形成聚集物,这与以前关于Lphn2与Ten3结合的报道相一致。
Lphn2与Ten3的结合有可能触发轴突内部信号通路的激活,从而导致轴突移向或远离发生相互作用的区域。由于表达Ten3的轴突和表达Lphn2的轴突所针对的区域并不重叠,Pederick及其同事们推断,Ten3-Lphn2的相互作用可能会导致排斥。为了测试这一点,这些作者使用了一种巧妙的方法,即把经过基因改造的病毒注入海马体,以操纵CA1神经元和及其下托目标区域的神经元中的Lphn2和Ten3表达。
这些作者将一种表达Lphn2的病毒注射到发育中的下托的远端(那里的Lphn2水平通常较低),以提高那里的Lphn2水平。一旦海马体发育完全,他们将一种表达荧光蛋白的病毒注射到Ten3表达较高的CA1近端,以可视化观察来自该区域的支配下托的轴突。这些轴突避开了人工表达Lphn2的区域。他们随后进行了相反的实验:减少Lphn2在发育中的近端下托的表达,这是因为该蛋白在那里的表达通常很高。在这种情况下,来自近端CA1区域的表达Ten3的轴突侵入了Lphn2表达减少的区域。这些结果表明,Lphn2与Ten3的相互作用对表达Ten3的轴突的排斥性引导是必要和充分的。
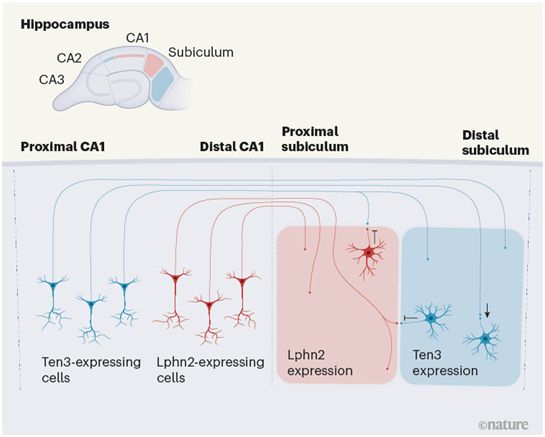
吸引性和排斥性的相互作用介导了神经回路的组装,图片来自Nature, 2021, doi:10.1038/d41586-021-01502-0。
重要的是,这些作者评估了Ten3介导的对表达Ten3的近端CA1轴突的吸引和Lphn2介导的对表达Ten3的近端CA1轴突的排斥的相对贡献,为此,他们减少整个下托中的Ten3和Lphn2的表达。这种基因操纵导致了非目标区域中的轴突增加和目标区域中的神经支配的减少。因此,表达Ten3的CA1近端轴突的精确靶向似乎需要Lphn2介导的远离非目标区域的排斥和Ten3介导的吸引。
表达Lphn2的CA1轴突对含有高水平Ten3的目标区域如何作出反应?与对照组小鼠的海马体相比,剔除远端下托细胞的Ten3表达导致表达Lphn2的轴突从CA1的较远部位进入下托的更远端,这表明来自目标区域的Ten3会排斥表达Lphn2的轴突。
轴突的吸引和排斥之间的合作是神经回路发育中一个熟悉的主题。这项研究表明,海马体神经回路网络中CA1轴突和下托神经元之间的连接严格遵循“Ten3轴突到Ten3目标区域,Lphn2轴突到Lphn2目标区域”的规则,这是由表达Ten3的细胞与表达Lphn2的细胞之间的相互排斥所决定的。
Pederick等人漂亮地展示了细胞表面分子之间的结合作用如何取决于细胞背景:在CA1轴突上,Ten3作为吸引性(Ten3)和排斥性(Lphn2)的目标区域来源线索的受体,而在下托,它的作用是排斥表达Lphn2的轴突。早前的一项研究已表明,Lphn2、Ten3和另一种细胞表面分子Flrt2的同时结合是CA1神经元与它们在海马体神经回路上游的搭档之间形成神经元突触连接所必需的,而不是用于轴突引导,这进一步说明了发育背景对指导神经回路组装的重要性。
还需要开展进一步的研究来确定由细胞表面分子(如Ten3和Lphn2)触发的信号级联反应,并确定轴突是否以及如何被一种特定的分子线索吸引或排斥。此外,如果吸引和排斥对精确的神经回路组装都是必要的,那么介导表达Lphn2的轴突的吸引的细胞表面分子的身份是什么?
基因组编码的细胞表面分子数量有限,但哺乳动物大脑的神经回路却非常复杂,每个参与引导轴突到其适当目标区域的细胞表面分子可能在不同的神经回路中发挥多种这样的功能,这取决于细胞背景和发育背景。在不同的神经回路的组装过程中,阐明每种细胞表面分子的背景依赖性作用将是至关重要的。(生物谷 Bioon.com)
参考资料:
Daniel T. Pederick et al. Reciprocal repulsions instruct the precise assembly of parallel hippocampal networks. Science, 2021, doi:10.1126/science.abg1774.
Yajun Xie et al. Attraction and repulsion cooperate during brain-circuit wiring. Nature, 2021, doi:10.1038/d41586-021-01502-0.
版权声明 本网站所有注明“来源:生物谷”或“来源:bioon”的文字、图片和音视频资料,版权均属于生物谷网站所有。非经授权,任何媒体、网站或个人不得转载,否则将追究法律责任。取得书面授权转载时,须注明“来源:生物谷”。其它来源的文章系转载文章,本网所有转载文章系出于传递更多信息之目的,转载内容不代表本站立场。不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
