
关于线粒体的重要作用,这些研究值得一读!
来源:本站原创 2019-10-21 01:22
2019年10月21日 讯 /生物谷BIOON/ --本期为大家带来的是线粒体与机体健康相关领域的最新研究进展,希望读者朋友能够喜欢。1. Ebiomedicine:线粒体调节因子或为癌症治疗新靶点DOI: 10.1016/j.ebiom.2019.09.017最近,来自Wistar研究所的研究人员发现了线粒体裂变因子(MFF)在控制癌细胞存活中的作用,表明该蛋白可能代表了有希望的治疗靶标。他们还
2019年10月21日 讯 /生物谷BIOON/ --本期为大家带来的是线粒体与机体健康相关领域的最新研究进展,希望读者朋友能够喜欢。
1. Ebiomedicine:线粒体调节因子或为癌症治疗新靶点
DOI: 10.1016/j.ebiom.2019.09.017

最近,来自Wistar研究所的研究人员发现了线粒体裂变因子(MFF)在控制癌细胞存活中的作用,表明该蛋白可能代表了有希望的治疗靶标。他们还发现,MFF的表达受到Myc的调控。这些结果在线发表在《EBioMedicine》杂志上。
线粒体是向我们的细胞提供能量的细胞器,它还控制多种细胞死亡机制,并在癌症中扮演着复杂的角色。此外,线粒体动力学能够协调细胞内线粒体大小,形状和位置,进而影响肿瘤的进展,但是直到现在,其机制还没有得到完整的阐明。
研究的资深作者, Wistar总裁兼首席执行官,癌症研究中心主任, Dario C. Altieri教授说:“我们知道,线粒体功能的重编程对于癌症的发展和转移至关重要。我们的发现揭示了这一过程中的新参与者和新途径,为选择性消除患者肿瘤细胞打开了具体的治疗机会。”
Altieri的实验室和一个国际合作团队表明,MFF基因在前列腺癌患者中扩增,与疾病复发和生存率降低相关。他们还观察到与正常组织相比,MFF蛋白在前列腺癌的小鼠模型以及患有其他癌症类型(包括肺癌和多发性骨髓瘤)的患者的组织样品中的表达升高。
重要的是,研究人员认为Myc癌蛋白是MFF表达的主要驱动力,它通常在多种癌症类型中扩增并控制肿瘤进展过程中的线粒体重编程。
Altieri及其同事表明,在癌症中,MFF与细胞死亡的线粒体调节剂VDAC1相互作用,从而关闭了其保持肿瘤细胞存活的功能。研究人员发现,在临床前模型中,MFF-VDAC1复合物的破坏激活了线粒体细胞死亡的多种机制,抑制了肿瘤细胞的增殖并降低了肿瘤的生长。
2. Circulation:新研究揭示调节线粒体压力反应的新靶点
DOI: 10.1161/CIRCULATIONAHA.118.037968

心脏的应激反应蛋白在心脏病发作时会被激活,以帮助防止细胞死亡。对此,坦普尔大学的Lewis Katz医学院研究人员首次表明,这些被称为MCUB的专门性应急蛋白,可暂时降低钙转运到线粒体的水平,从而缓解心脏病的严重程度。相关结果于近日发表在《Circulation》杂志上。
该研究表明,MCUB可以作为研究和治疗以钙超载和细胞死亡为特征的疾病(包括心力衰竭,心脏病发作,中风和神经退行性疾病)的有希望的新靶标。
“ MCUB调节受损心脏组织中线粒体的钙吸收,从而限制钙超载的发生。众所周知,钙超载是导致细胞死亡(尤其是心脏病发作后)的主要因素,”该研究副教授John W. Elrod博士解释说。
钙稳态对许多日常细胞活动至关重要,线粒体是调节调节钙稳态的主要部位。为了使钙进入线粒体,钙离子需要穿过称为线粒体钙单向转运蛋白(MCU)的,位于线粒体内膜上的通道。线粒体吸收的钙离子水平受该通道各种成分的调节。此外,尽管MCUB与亚基构成MCU非常相似,但前者在钙调节中的确切作用仍是未知的。
新的的研究发现,细胞中MCUB基因的缺失会导致构成钙通道的蛋白质发生变化,这对于控制该通道的开启或关闭至关重要。由于这些改变是由应激(例如心脏细胞损伤)诱导的,因此研究人员接下来研究了小鼠心脏病发作后MCUB的作用。在遭受心脏病发作的小鼠中,研究小组观察到MCUB基因表达显着升高,而MCU和通道MICU1表达水平则有所降低。当在小鼠中引起心脏病发作之前进行基因表达时,MCUB改变了通道以减少受伤心脏中的钙超载,最终减少了组织损伤。
Elrod的团队还发现,尽管可以改善心脏损伤后的细胞存活率,但增加的MCUB活性却以线粒体能量产生为代价。Elrod博士解释说:“ MCUB的引入是一种补偿性变化。”
“ MCUB为我们提供了一个新的分子靶标进行研究。它的独特之处在于它改变了通道的化学计量,从而提出了一种新的机制,可以对治疗操作进行修正。我们认为调节MCUB可以使我们在不完全抑制所有能量功能的情况下降低线粒体钙的吸收。”
最后,作者希望通过后续研究确定分子相互作用的确切位置,可以为如何针对心脏病中的线粒体钙超负荷提供更多见解。
3. PNAS:失控的线粒体会引起细胞端粒损伤
doi/10.1073/pnas.1910574116
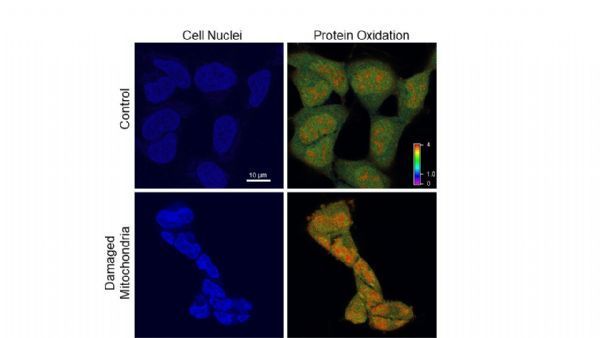
匹兹堡大学希尔曼癌症中心的研究人员为长期以来的观点提供了第一个具体证据,即患病的线粒体污染了它们本应提供能量的细胞。
这篇近日发表在《PNAS》上的论文涉及一项因果实验,目的是启动线粒体连锁反应,这种反应会对细胞造成破坏,一直到遗传水平。
匹兹堡大学医学院和希尔曼癌症中心的药理学和化学生物学教授、资深作家Bennett Van Houten博士说:"我喜欢把它叫做'切尔诺贝利效应'--你已经打开了反应堆,现在却不能关掉它。这台清洁燃烧的机器现在正在疯狂地污染环境,而这种污染会反过来损害电子传输功能。这是一个恶性循环。"
Van Houten的团队使用了一项由卡内基梅隆大学的Marcel Bruchez博士发明的新技术,当暴露在光线下时,线粒体内会产生破坏性的活性氧物种,在本例中是单线态氧。
Van Houten说:"这就是切尔诺贝利事故。一旦你把灯关掉,就再也没有单线态氧了,但是你破坏了电子传递链,所以48小时后,线粒体仍然在释放活性氧--但是细胞没有死亡,它们只是坐在那里爆发。"
此时,细胞核受到自由基的冲击。它会收缩和扭曲。细胞停止分裂。然而,DNA似乎出奇地完整。
"如果你把染色体想象成一辆车,端粒的宽度就是车牌的宽度,"匹兹堡大学公共卫生研究生院和希尔曼大学环境与职业健康教授Patricia Opresko博士说。
因此,为了研究线粒体熔毁的遗传效应,研究人员不得不用荧光标记点亮那些微小的端盖,结果他们发现了端粒脆弱和断裂的明显迹象。
然后,在关键的一步,研究人员在线粒体失活的细胞上重复了整个实验。没有线粒体来维持反应,细胞内就没有自由基的积累,也没有端粒的损伤。
"基本上,我们在机器有机会造成任何损害之前就把它关掉了,"Van Houten说。
Van Houten建议,这些发现可以用于改善光动力癌症治疗,包括使用光纤传输的光用活性氧轰击实体肿瘤。
他的团队在这些实验中发现的一件事是,抑制ATM(一种信号DNA损伤的蛋白质)会放大线粒体释放出的活性氧的破坏作用。细胞不仅萎缩,而且死亡。
Van Houten说,通过将光动力疗法与ATM抑制相结合,可能设计出一种利用光有效杀死癌细胞的系统。
4. Nature:在线粒体中鉴定出一种ATP敏感性的钾离子通道
doi:10.1038/s41586-019-1498-3.
线粒体是一种特殊的细胞器,这是因为它们具有自己的DNA,称为线粒体DNA(mtDNA)。与存在于细胞核中的更大的DNA集合(基因组)不同的是,mtDNA仅通过母体的卵细胞进行传递。
mtDNA也比细胞核DNA更容易在它的DNA代码中产生随机变化或突变,这些变化或变异随着人的年龄增长而增加,不过,这种情形也会在生殖细胞的发育过程中发生,从而导致遗传性疾病。在美国儿童中,这些遗传性疾病的发病率大约为1/4300。
线粒体是我们细胞的能量工厂。每天,人类需要身体产生ATP来为所有细胞活动提供能量。神经冲动、肌肉收缩、DNA复制和蛋白合成仅是依赖于ATP供应的至关重要的过程的一些例子。
线粒体以ATP的形式为内源性反应提供化学能,它们的活性必须满足细胞能量需求,但是将这种细胞器性能与ATP水平相关联在一起的机制却知之甚少。
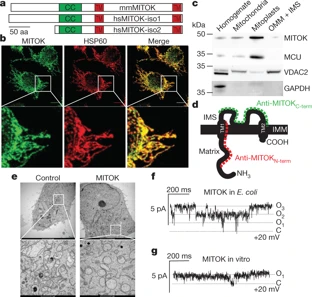
在一项新的研究中,来自意大利帕多瓦大学的研究人员证实一种存在于线粒体中的蛋白复合物介导ATP依赖性钾电流,这种蛋白复合物称为mitoKATP。相关研究结果近期发表在Nature期刊上,论文标题为“Identification of an ATP-sensitive potassium channel in mitochondria”。
这些研究人员发现钾离子通道mitoKATP类似于它们在细胞膜上的对应物,由成孔亚基和ATP结合亚基组成,他们将这两个亚基分别称之为MITOK和MITOSUR。对MITOK和MITOSUR的体外重建概括了mitoKATP的主要特性。
MITOK的过度表达引发显著的细胞器肿胀,然而通过遗传手段剔除这个亚基会导致线粒体膜电位的不稳定性,嵴内腔 (intracristal space)扩大和氧化磷酸化减少。在小鼠模型中,MITOK的缺失抑制了由重氮氧化合物(diazoxide)诱导的药物预处理引起的心脏保护作用。
这些研究结果表明,mitoKATP通道通过调节细胞器的体积和功能来响应细胞的能量状态,从而在线粒体生理学和对几种病理过程的潜在影响中起关键作用。
5. Nature:揭示H+转运是线粒体ADP/ATP载体发挥功能所必需的一种功能
doi:10.1038/s41586-019-1400-3.
一种称为线粒体的亚细胞结构是我们细胞的能量工厂。每天,人类需要身体产生ATP来为所有细胞活动提供能量。神经冲动、肌肉收缩、DNA复制和蛋白合成仅是依赖于ATP供应的至关重要的过程的一些例子。
鉴于我们体内仅含有少量的ATP,我们需要使用位于线粒体中的一种称为ATP合酶(ATP synthase)的酶复合物,将ATP降解时产生的产物ADP(二磷酸腺苷)和磷酸盐重新转化为ATP。通过这种方式,每个ATP分子每天大约循环回收1300次。为了让ADP到达ATP合酶,并让重新产生的ATP为细胞提供能量,每个ADP/ATP分子必须穿过包围着线粒体的不可渗透的脂质膜。线粒体ADP/ATP载体(mitochondrial ADP/ATP carrier, AAC)参与线粒体中的ADP和ATP转运。
AAC在两种状态之间循环:在一种称为细胞质开放状态(cytoplasmic-open state)的状态下,它的中心结合位点可用于结合ADP,而在另一种称为基质开放状态(matrix-open state)的状态下,这种结合位点可用于结合新合成的ATP。
AAC是线粒体内膜的主要转运蛋白。它利用线粒体中的ATP交换细胞质中的ADP,并且控制细胞中的ATP产生。此外,人们已提出AAC介导线粒体解偶联(mitochondrial uncoupling),但是却难以证实这种功能或阐明它的作用机制。
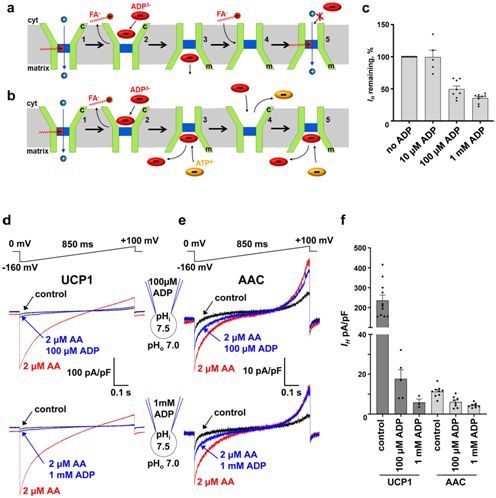
在一项新的研究中,来自美国加州大学旧金山分校和哈佛医学院等研究机构的研究人员记录了直接来自各种小鼠组织的线粒体内膜的AAC电流,并鉴定出两种不同的转运模式:ADP/ATP交换和氢离子(H+)转运。相关研究结果发表在2019年7月25日的Nature期刊上,论文标题为“H+ transport is an integral function of the mitochondrial ADP/ATP carrier”。
AAC介导的H+电流需要游离脂肪酸,并且类似于通过在棕色脂肪中发现的产热解偶联蛋白1(uncoupling protein 1)发生的H+泄漏。AAC的ADP/ATP交换负向调节H+泄漏,但并没有完全抑制它。这表明H+泄漏和线粒体解偶联可能通过细胞的ATP需求和ADP/ATP交换速率来加以动态控制。
通过介导这两种不同的转运模式:ADP/ATP交换和H+泄漏,AAC将线粒体中的偶联能量转换(ATP产生)和解偶联能量转换(产热)相关联在一起。
6. Cancer Discovery:KRAS诱导线粒体自噬来促进胰腺癌发展
DOI: 10.1158/2159-8290.CD-18-1409
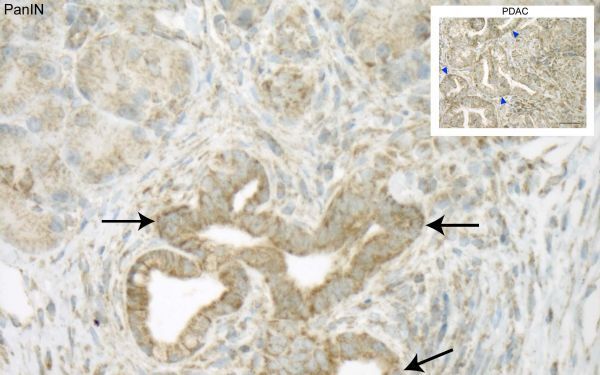
癌细胞在肿瘤的低能量环境中使用一种奇怪的繁殖策略:他们破坏了自己的线粒体!冷泉港实验室(CSHL)的研究人员现在也知道了这个过程是如何发生的,为胰腺癌治疗提供了一个有希望的新靶点。
为什么癌细胞想要破坏自己的线粒体功能?医学博士Brinda Alagesan承认:"这似乎非常违反直觉。"她是CSHL David Tuveson博士实验室的成员。
根据Alagesan的观点,要想知道癌细胞为什么会这样做,最简单的方法就是把线粒体看作动力装置。"线粒体是细胞的动力源,"她说道。就像传统的动力装置一样,线粒体也会产生污染。
Alagesan补充说:"这些有害的副产品或污染物被称为活性氧(ROS)。"其中很多会对细胞造成损害。我们相信,胰腺癌细胞(通过吞噬自身的线粒体)正在减少这些破坏性活性氧的产生,同时仍在制造足够的能量进行增殖。"
这仍然是一个假设,但它可以解释为什么胰腺癌细胞更容易发生自噬,一种形式的自噬或"自食"线粒体。
在《Cancer Discovery》杂志上,Alagesan和共同的主要作者Timothy Humpton博士描述了当一种叫做KRAS的蛋白质在胰腺肿瘤独特的营养缺乏的环境中变得活跃时会发生什么。KRAS启动了一个"信号级联",导致细胞吞噬自己的线粒体,葡萄糖和谷氨酰胺从剩余的线粒体转移。这些转移的营养物质被用来支持细胞分裂。
Alagesan解释说:"理想情况下,我们想要直接抑制促进KRAS蛋白的癌症,但不幸的是,到目前为止还没有人能够以临床相关的方式做到这一点。"
Tuveson团队没有直接阻止KRAS,而是跟踪了KRAS激活后的蛋白信号级联。他们发现了一种导致蛋白质NIX增加的途径。NIX是触发线粒体自噬的直接原因,而线粒体自噬对于癌细胞的增殖是至关重要的。
Alagesan说:"小鼠实验结果向我们表明,通过抑制NIX通路,我们可以阻止癌细胞以它们增殖所需的方式利用能量。"
Tuveson团队现在将注意力转向阻断人类胰腺癌细胞中的NIX通路,并将其应用于临床试验的设计。
7. Nature:揭示线粒体质量控制缺陷可导致心脏病
doi:10.1038/s41586-019-1667-4.

一个编码腺嘌呤核苷酸转运蛋白(adenine nucleotide translocator, ANT)的基因中的突变会导致很多疾病,比如心脏病和眼肌无力,但是这些突变如何引发疾病的内在机制尚不清楚。如今,在一项新的研究中,来自美国宾夕法尼亚大学佩雷尔曼医学院的研究人员揭示了ANT的令人惊讶的新功能:ANT对于一种称为线粒体自噬(mitophagy)---通过清除受损的线粒体,有助于确保线粒体网络的完整性---的质量控制过程至关重要,并且发现导致这种质量控制系统出现缺陷的ANT突变最终会导致心脏病。相关研究结果近期发表在Nature期刊上,论文标题为“The ADP/ATP translocase drives mitophagy independent of nucleotide exchange”。
ANT是一种众所周知的蛋白,有助于线粒体产生驱动体内细胞正常运作所需的化学能,即三磷酸腺苷(ATP)。尽管已知ANT基因突变会引起疾病,包括心肌病(一种使人的心脏更难将血液泵送到身体其他部位的疾病),但是已有研究表明这些突变不会影响ANT产生化学能的能力,这引发了有关人们将如何患病的问题。
论文通讯作者、宾夕法尼亚大学佩雷尔曼医学院心血管医学教授Zoltan Arany博士说,“在揭示ANT与线粒体自噬之间的关联性以及ANT突变对线粒体质量控制的影响时,我们的发现改变了我们对这些致病突变的思考方式,这能够让我们将注意力集中在正确的途径上。如今,我们知道这些疾病是由线粒体质量控制缺陷而不是由ATP缺乏引起的,我们可以开始考虑改善线粒体质量控制的治疗方法。”
作为细胞的能量工厂,线粒体产生驱动体内细胞正常运作所需的大部分ATP,并且在包括心脏和肝脏在内的高能量需求的器官中发挥着重要作用。线粒体质量控制机制,包括线粒体自噬(当线粒体在遭受损伤或应激后出现缺陷时,通常就会发生),有助于确保线粒体网络正常运行。尽管已确定了线粒体自噬的作用,但是到目前为止,关于参与线粒体自噬的不同蛋白的信息仍然是有限的。
为了更深入地了解参与线粒体自噬的组分,这些研究人员使用了多个报告系统和促线粒体自噬触发因子来开展CRISPR/Cas9全基因组筛选。令他们吃惊的是,他们发现ANT是许多细胞类型中线粒体质量控制途径所必需的。在证实他们的发现的后续测试中,他们发现ANT促进了线粒体自噬,但这与它在产生ATP中的作用无关,这表明这种蛋白具有两种不同的功能。
基于这一发现,这些研究人员设计出研究ANT基因突变是否影响线粒体自噬的测试方法。他们发现,ANT中的致病突变清除了它与将蛋白从线粒体内膜重定位到线粒体基质中所必需的蛋白复合物结合的能力,从而最终抑制了线粒体自噬。进一步的研究表明通过遗传手段剔除ANT可以抑制线粒体自噬。他们还发现,缺乏ANT的小鼠具有较弱的线粒体自噬能力,因而经历了非典型线粒体的大量积累。
Arany说,“我们非常吃惊地发现ANT参与线粒体自噬,这揭示出它具有两种独立的功能---ATP产生和线粒体质量控制。尽管目前没有改善线粒体质量控制的灵丹妙药,但是我们希望我们的发现为今后的研究工作打下基础,从而有助于研究如何抵消对线粒体自噬的抑制,并且最终阻止这些突变引起疾病。”
8. Nature:揭示一种阻止蛋白聚集物在线粒体中聚集的新机制
doi:10.1038/s41586-019-1307-z.

蛋白聚集物对线粒体功能是有害的,因而会破坏向它们的宿主细胞提供化学能。在一项新的研究中,来自德国慕尼黑大学等研究机构的研究人员描述了一种阻止这些蛋白聚集物在线粒体中聚集的蛋白复合物。相关研究结果近期发表在Nature期刊上,论文标题为“Structure and function of Vms1 and Arb1 in RQC and mitochondrial proteome homeostasis”。
作为一种细胞器,线粒体为高等生物中的细胞提供它们的代谢和功能维持所必需的化学能。此外,许多必需代谢物的生物合成发生在这些细胞器中。因此,必须快速检测和修复它们的功能出现的任何扰动---比如,错误折叠蛋白的异常堆积。
在慕尼黑大学生物医学中心的Walter Neupert、慕尼黑大学基因中心的Roland Beckmann和日本仙台大学的Toshifumi Inada的领导下,这些研究人员如今阐明了细胞抑制前往线粒体的毒性蛋白聚集物形成的机制之一。如果不加以抑制的话,这些蛋白聚集物就会切断为不可或缺的细胞功能提供的能量。
蛋白由称为核糖体的细胞器合成。这些RNA-蛋白复合物(即核糖体)对从细胞核输出的确定着特定蛋白氨基酸序列的信使RNA(mRNA)蓝图进行解码。核糖体从确定的起始位点开始依次附着于mRNA分子上。这允许每个mRNA分子以装配线方式编程每种特定蛋白的许多拷贝的合成。如果这个过程受到中断而且前面的核糖体发生停滞,那么后面的核糖体堆积在后面。在这些条件下,它们附着的不完整蛋白(译者注:指的是因核糖体停滞,核糖体上的蛋白未完整合成)能够容易地彼此相互作用以形成蛋白聚集物。为解决此类交通拥堵,细胞激活“核糖体相关质量控制(ribosome-associated quality control, RQC)机制”。RQC途径的作用机制是让停滞的核糖体的两个亚基(60S大亚基和40S小亚基)分离开来(从而释放mRNA)并且将丙氨酸(A)和苏氨酸(T)附着到60S亚基上不完整蛋白的停滞羧基末端(译者注:指的是因核糖体停滞,核糖体在mRNA的特定局部大量堆积,核糖体上的新生蛋白未能完整合成,这就意味着在发生核糖体停滞后,新生蛋白的羧基末端停滞了,也就是羧基末端不再添加新的氨基酸)上。这样,这种不完整的蛋白从核糖体中释放出来,“CAT尾巴”标志着它随后会被降解。
然而,就线粒体蛋白而言,情况就更加复杂了。大多数受到抑制或发生停滞的线粒体蛋白是由附着在线粒体膜孔上的核糖体在细胞质中形成的,这些新生的线粒体蛋白通过这些孔直接进入这种细胞器。这种合成与线粒体摄取的紧密连接阻碍了CAT标记的蛋白在细胞质中的释放和裂解。此外,CAT标记的蛋白本身具有增加的聚集倾向,因此它们的输入对线粒体功能尤其有害。
幸运的是,细胞已进化出一种专门用于线粒体蛋白的RQC替代形式。这项新研究的作者之前已证实Vms1蛋白在这一途径中发挥着关键作用,但这种RQC模式的确切机制尚不清楚。Beckmann说,“通过将基于低温电镜的结构分析与生化、分子生物学和遗传学实验相结合,我们如今成功地阐明了Vms1的工作机制。”Vms1能够解离核糖体并释放出停滞的蛋白,不论这种停滞的蛋白是否携带CAT序列。因此,它有效地抵消了添加到线粒体蛋白上的CAT序列,从而降低了蛋白聚集的风险以及对线粒体和细胞功能的不利影响。此外,Beckmann及其同事们发现了一种之前未被描述的在这整个过程中发挥作用的蛋白:Arb1,并展示了它与Vms1之间如何相互作用。这些新的研究结果可能有助于更好地了解各种疾病,包括代谢疾病和神经退行性疾病,这些疾病与线粒体功能的损害有关。
9. Nature:线粒体裂殖需要蛋白DRP1,但不需要动力蛋白
doi:10.1038/s41586-019-1296-y.
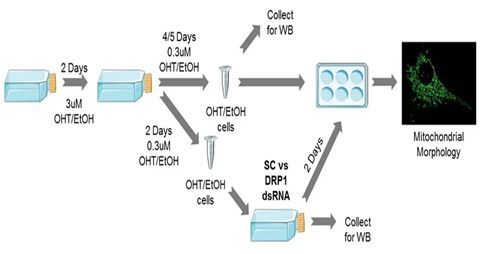
线粒体裂变(mitochondrial fission,有时也译作线粒体分裂)是维持线粒体网络所必需的,并且依赖于一种称为动力蛋白相关蛋白1(dynamin-relatedprotein 1, DRP1,也称为DNM1L)的GTP酶。DRP1形成螺旋寡聚体,包裹线粒体外膜并将其分裂。最近,有人提出DRP1不足以进行线粒体裂变,另一种称为动力蛋白-2(dynamin-2, DNM2, 也称为DYN2)的GTP酶是线粒体分裂复合物的重要组成部分。
在一项新的研究中,来自德国哥廷根大学医学中心的研究人员报道缺乏所有三种哺乳动物动力蛋白(DNM1、DNM2和DNM3)的小鼠成纤维细胞(下称动力蛋白三重敲除细胞)以及仅敲降DNM2的细胞既没有表现出线粒体裂变或过度融合缺陷,但是敲降DRP1可很容易检测到这些缺陷,即便在动力蛋白三重敲除细胞中,也是如此。相关研究结果发表在2019年6月20日的Nature期刊上,论文标题为“Mitochondrial fission requires DRP1 but not dynamins”。
鉴于线粒体和过氧化物酶体具有相同的裂变机制,这些研究人员研究了过氧化物酶体裂变,结果观察到与线粒体裂变相同的情形。
因此,这些研究结果表明DRP1对于线粒体和过氧化物酶体裂变是必需的,而哺乳动物动力蛋白DNM1、DNM2和DNM3则并不是必需的。(生物谷Bioon.com)
1. Ebiomedicine:线粒体调节因子或为癌症治疗新靶点
DOI: 10.1016/j.ebiom.2019.09.017
最近,来自Wistar研究所的研究人员发现了线粒体裂变因子(MFF)在控制癌细胞存活中的作用,表明该蛋白可能代表了有希望的治疗靶标。他们还发现,MFF的表达受到Myc的调控。这些结果在线发表在《EBioMedicine》杂志上。
线粒体是向我们的细胞提供能量的细胞器,它还控制多种细胞死亡机制,并在癌症中扮演着复杂的角色。此外,线粒体动力学能够协调细胞内线粒体大小,形状和位置,进而影响肿瘤的进展,但是直到现在,其机制还没有得到完整的阐明。
研究的资深作者, Wistar总裁兼首席执行官,癌症研究中心主任, Dario C. Altieri教授说:“我们知道,线粒体功能的重编程对于癌症的发展和转移至关重要。我们的发现揭示了这一过程中的新参与者和新途径,为选择性消除患者肿瘤细胞打开了具体的治疗机会。”
Altieri的实验室和一个国际合作团队表明,MFF基因在前列腺癌患者中扩增,与疾病复发和生存率降低相关。他们还观察到与正常组织相比,MFF蛋白在前列腺癌的小鼠模型以及患有其他癌症类型(包括肺癌和多发性骨髓瘤)的患者的组织样品中的表达升高。
重要的是,研究人员认为Myc癌蛋白是MFF表达的主要驱动力,它通常在多种癌症类型中扩增并控制肿瘤进展过程中的线粒体重编程。
Altieri及其同事表明,在癌症中,MFF与细胞死亡的线粒体调节剂VDAC1相互作用,从而关闭了其保持肿瘤细胞存活的功能。研究人员发现,在临床前模型中,MFF-VDAC1复合物的破坏激活了线粒体细胞死亡的多种机制,抑制了肿瘤细胞的增殖并降低了肿瘤的生长。
2. Circulation:新研究揭示调节线粒体压力反应的新靶点
DOI: 10.1161/CIRCULATIONAHA.118.037968
心脏的应激反应蛋白在心脏病发作时会被激活,以帮助防止细胞死亡。对此,坦普尔大学的Lewis Katz医学院研究人员首次表明,这些被称为MCUB的专门性应急蛋白,可暂时降低钙转运到线粒体的水平,从而缓解心脏病的严重程度。相关结果于近日发表在《Circulation》杂志上。
该研究表明,MCUB可以作为研究和治疗以钙超载和细胞死亡为特征的疾病(包括心力衰竭,心脏病发作,中风和神经退行性疾病)的有希望的新靶标。
“ MCUB调节受损心脏组织中线粒体的钙吸收,从而限制钙超载的发生。众所周知,钙超载是导致细胞死亡(尤其是心脏病发作后)的主要因素,”该研究副教授John W. Elrod博士解释说。
钙稳态对许多日常细胞活动至关重要,线粒体是调节调节钙稳态的主要部位。为了使钙进入线粒体,钙离子需要穿过称为线粒体钙单向转运蛋白(MCU)的,位于线粒体内膜上的通道。线粒体吸收的钙离子水平受该通道各种成分的调节。此外,尽管MCUB与亚基构成MCU非常相似,但前者在钙调节中的确切作用仍是未知的。
新的的研究发现,细胞中MCUB基因的缺失会导致构成钙通道的蛋白质发生变化,这对于控制该通道的开启或关闭至关重要。由于这些改变是由应激(例如心脏细胞损伤)诱导的,因此研究人员接下来研究了小鼠心脏病发作后MCUB的作用。在遭受心脏病发作的小鼠中,研究小组观察到MCUB基因表达显着升高,而MCU和通道MICU1表达水平则有所降低。当在小鼠中引起心脏病发作之前进行基因表达时,MCUB改变了通道以减少受伤心脏中的钙超载,最终减少了组织损伤。
Elrod的团队还发现,尽管可以改善心脏损伤后的细胞存活率,但增加的MCUB活性却以线粒体能量产生为代价。Elrod博士解释说:“ MCUB的引入是一种补偿性变化。”
“ MCUB为我们提供了一个新的分子靶标进行研究。它的独特之处在于它改变了通道的化学计量,从而提出了一种新的机制,可以对治疗操作进行修正。我们认为调节MCUB可以使我们在不完全抑制所有能量功能的情况下降低线粒体钙的吸收。”
最后,作者希望通过后续研究确定分子相互作用的确切位置,可以为如何针对心脏病中的线粒体钙超负荷提供更多见解。
3. PNAS:失控的线粒体会引起细胞端粒损伤
doi/10.1073/pnas.1910574116
匹兹堡大学希尔曼癌症中心的研究人员为长期以来的观点提供了第一个具体证据,即患病的线粒体污染了它们本应提供能量的细胞。
这篇近日发表在《PNAS》上的论文涉及一项因果实验,目的是启动线粒体连锁反应,这种反应会对细胞造成破坏,一直到遗传水平。
匹兹堡大学医学院和希尔曼癌症中心的药理学和化学生物学教授、资深作家Bennett Van Houten博士说:"我喜欢把它叫做'切尔诺贝利效应'--你已经打开了反应堆,现在却不能关掉它。这台清洁燃烧的机器现在正在疯狂地污染环境,而这种污染会反过来损害电子传输功能。这是一个恶性循环。"
Van Houten的团队使用了一项由卡内基梅隆大学的Marcel Bruchez博士发明的新技术,当暴露在光线下时,线粒体内会产生破坏性的活性氧物种,在本例中是单线态氧。
Van Houten说:"这就是切尔诺贝利事故。一旦你把灯关掉,就再也没有单线态氧了,但是你破坏了电子传递链,所以48小时后,线粒体仍然在释放活性氧--但是细胞没有死亡,它们只是坐在那里爆发。"
此时,细胞核受到自由基的冲击。它会收缩和扭曲。细胞停止分裂。然而,DNA似乎出奇地完整。
"如果你把染色体想象成一辆车,端粒的宽度就是车牌的宽度,"匹兹堡大学公共卫生研究生院和希尔曼大学环境与职业健康教授Patricia Opresko博士说。
因此,为了研究线粒体熔毁的遗传效应,研究人员不得不用荧光标记点亮那些微小的端盖,结果他们发现了端粒脆弱和断裂的明显迹象。
然后,在关键的一步,研究人员在线粒体失活的细胞上重复了整个实验。没有线粒体来维持反应,细胞内就没有自由基的积累,也没有端粒的损伤。
"基本上,我们在机器有机会造成任何损害之前就把它关掉了,"Van Houten说。
Van Houten建议,这些发现可以用于改善光动力癌症治疗,包括使用光纤传输的光用活性氧轰击实体肿瘤。
他的团队在这些实验中发现的一件事是,抑制ATM(一种信号DNA损伤的蛋白质)会放大线粒体释放出的活性氧的破坏作用。细胞不仅萎缩,而且死亡。
Van Houten说,通过将光动力疗法与ATM抑制相结合,可能设计出一种利用光有效杀死癌细胞的系统。
4. Nature:在线粒体中鉴定出一种ATP敏感性的钾离子通道
doi:10.1038/s41586-019-1498-3.
线粒体是一种特殊的细胞器,这是因为它们具有自己的DNA,称为线粒体DNA(mtDNA)。与存在于细胞核中的更大的DNA集合(基因组)不同的是,mtDNA仅通过母体的卵细胞进行传递。
mtDNA也比细胞核DNA更容易在它的DNA代码中产生随机变化或突变,这些变化或变异随着人的年龄增长而增加,不过,这种情形也会在生殖细胞的发育过程中发生,从而导致遗传性疾病。在美国儿童中,这些遗传性疾病的发病率大约为1/4300。
线粒体是我们细胞的能量工厂。每天,人类需要身体产生ATP来为所有细胞活动提供能量。神经冲动、肌肉收缩、DNA复制和蛋白合成仅是依赖于ATP供应的至关重要的过程的一些例子。
线粒体以ATP的形式为内源性反应提供化学能,它们的活性必须满足细胞能量需求,但是将这种细胞器性能与ATP水平相关联在一起的机制却知之甚少。
在一项新的研究中,来自意大利帕多瓦大学的研究人员证实一种存在于线粒体中的蛋白复合物介导ATP依赖性钾电流,这种蛋白复合物称为mitoKATP。相关研究结果近期发表在Nature期刊上,论文标题为“Identification of an ATP-sensitive potassium channel in mitochondria”。
这些研究人员发现钾离子通道mitoKATP类似于它们在细胞膜上的对应物,由成孔亚基和ATP结合亚基组成,他们将这两个亚基分别称之为MITOK和MITOSUR。对MITOK和MITOSUR的体外重建概括了mitoKATP的主要特性。
MITOK的过度表达引发显著的细胞器肿胀,然而通过遗传手段剔除这个亚基会导致线粒体膜电位的不稳定性,嵴内腔 (intracristal space)扩大和氧化磷酸化减少。在小鼠模型中,MITOK的缺失抑制了由重氮氧化合物(diazoxide)诱导的药物预处理引起的心脏保护作用。
这些研究结果表明,mitoKATP通道通过调节细胞器的体积和功能来响应细胞的能量状态,从而在线粒体生理学和对几种病理过程的潜在影响中起关键作用。
5. Nature:揭示H+转运是线粒体ADP/ATP载体发挥功能所必需的一种功能
doi:10.1038/s41586-019-1400-3.
一种称为线粒体的亚细胞结构是我们细胞的能量工厂。每天,人类需要身体产生ATP来为所有细胞活动提供能量。神经冲动、肌肉收缩、DNA复制和蛋白合成仅是依赖于ATP供应的至关重要的过程的一些例子。
鉴于我们体内仅含有少量的ATP,我们需要使用位于线粒体中的一种称为ATP合酶(ATP synthase)的酶复合物,将ATP降解时产生的产物ADP(二磷酸腺苷)和磷酸盐重新转化为ATP。通过这种方式,每个ATP分子每天大约循环回收1300次。为了让ADP到达ATP合酶,并让重新产生的ATP为细胞提供能量,每个ADP/ATP分子必须穿过包围着线粒体的不可渗透的脂质膜。线粒体ADP/ATP载体(mitochondrial ADP/ATP carrier, AAC)参与线粒体中的ADP和ATP转运。
AAC在两种状态之间循环:在一种称为细胞质开放状态(cytoplasmic-open state)的状态下,它的中心结合位点可用于结合ADP,而在另一种称为基质开放状态(matrix-open state)的状态下,这种结合位点可用于结合新合成的ATP。
AAC是线粒体内膜的主要转运蛋白。它利用线粒体中的ATP交换细胞质中的ADP,并且控制细胞中的ATP产生。此外,人们已提出AAC介导线粒体解偶联(mitochondrial uncoupling),但是却难以证实这种功能或阐明它的作用机制。
在一项新的研究中,来自美国加州大学旧金山分校和哈佛医学院等研究机构的研究人员记录了直接来自各种小鼠组织的线粒体内膜的AAC电流,并鉴定出两种不同的转运模式:ADP/ATP交换和氢离子(H+)转运。相关研究结果发表在2019年7月25日的Nature期刊上,论文标题为“H+ transport is an integral function of the mitochondrial ADP/ATP carrier”。
AAC介导的H+电流需要游离脂肪酸,并且类似于通过在棕色脂肪中发现的产热解偶联蛋白1(uncoupling protein 1)发生的H+泄漏。AAC的ADP/ATP交换负向调节H+泄漏,但并没有完全抑制它。这表明H+泄漏和线粒体解偶联可能通过细胞的ATP需求和ADP/ATP交换速率来加以动态控制。
通过介导这两种不同的转运模式:ADP/ATP交换和H+泄漏,AAC将线粒体中的偶联能量转换(ATP产生)和解偶联能量转换(产热)相关联在一起。
6. Cancer Discovery:KRAS诱导线粒体自噬来促进胰腺癌发展
DOI: 10.1158/2159-8290.CD-18-1409
癌细胞在肿瘤的低能量环境中使用一种奇怪的繁殖策略:他们破坏了自己的线粒体!冷泉港实验室(CSHL)的研究人员现在也知道了这个过程是如何发生的,为胰腺癌治疗提供了一个有希望的新靶点。
为什么癌细胞想要破坏自己的线粒体功能?医学博士Brinda Alagesan承认:"这似乎非常违反直觉。"她是CSHL David Tuveson博士实验室的成员。
根据Alagesan的观点,要想知道癌细胞为什么会这样做,最简单的方法就是把线粒体看作动力装置。"线粒体是细胞的动力源,"她说道。就像传统的动力装置一样,线粒体也会产生污染。
Alagesan补充说:"这些有害的副产品或污染物被称为活性氧(ROS)。"其中很多会对细胞造成损害。我们相信,胰腺癌细胞(通过吞噬自身的线粒体)正在减少这些破坏性活性氧的产生,同时仍在制造足够的能量进行增殖。"
这仍然是一个假设,但它可以解释为什么胰腺癌细胞更容易发生自噬,一种形式的自噬或"自食"线粒体。
在《Cancer Discovery》杂志上,Alagesan和共同的主要作者Timothy Humpton博士描述了当一种叫做KRAS的蛋白质在胰腺肿瘤独特的营养缺乏的环境中变得活跃时会发生什么。KRAS启动了一个"信号级联",导致细胞吞噬自己的线粒体,葡萄糖和谷氨酰胺从剩余的线粒体转移。这些转移的营养物质被用来支持细胞分裂。
Alagesan解释说:"理想情况下,我们想要直接抑制促进KRAS蛋白的癌症,但不幸的是,到目前为止还没有人能够以临床相关的方式做到这一点。"
Tuveson团队没有直接阻止KRAS,而是跟踪了KRAS激活后的蛋白信号级联。他们发现了一种导致蛋白质NIX增加的途径。NIX是触发线粒体自噬的直接原因,而线粒体自噬对于癌细胞的增殖是至关重要的。
Alagesan说:"小鼠实验结果向我们表明,通过抑制NIX通路,我们可以阻止癌细胞以它们增殖所需的方式利用能量。"
Tuveson团队现在将注意力转向阻断人类胰腺癌细胞中的NIX通路,并将其应用于临床试验的设计。
7. Nature:揭示线粒体质量控制缺陷可导致心脏病
doi:10.1038/s41586-019-1667-4.
一个编码腺嘌呤核苷酸转运蛋白(adenine nucleotide translocator, ANT)的基因中的突变会导致很多疾病,比如心脏病和眼肌无力,但是这些突变如何引发疾病的内在机制尚不清楚。如今,在一项新的研究中,来自美国宾夕法尼亚大学佩雷尔曼医学院的研究人员揭示了ANT的令人惊讶的新功能:ANT对于一种称为线粒体自噬(mitophagy)---通过清除受损的线粒体,有助于确保线粒体网络的完整性---的质量控制过程至关重要,并且发现导致这种质量控制系统出现缺陷的ANT突变最终会导致心脏病。相关研究结果近期发表在Nature期刊上,论文标题为“The ADP/ATP translocase drives mitophagy independent of nucleotide exchange”。
ANT是一种众所周知的蛋白,有助于线粒体产生驱动体内细胞正常运作所需的化学能,即三磷酸腺苷(ATP)。尽管已知ANT基因突变会引起疾病,包括心肌病(一种使人的心脏更难将血液泵送到身体其他部位的疾病),但是已有研究表明这些突变不会影响ANT产生化学能的能力,这引发了有关人们将如何患病的问题。
论文通讯作者、宾夕法尼亚大学佩雷尔曼医学院心血管医学教授Zoltan Arany博士说,“在揭示ANT与线粒体自噬之间的关联性以及ANT突变对线粒体质量控制的影响时,我们的发现改变了我们对这些致病突变的思考方式,这能够让我们将注意力集中在正确的途径上。如今,我们知道这些疾病是由线粒体质量控制缺陷而不是由ATP缺乏引起的,我们可以开始考虑改善线粒体质量控制的治疗方法。”
作为细胞的能量工厂,线粒体产生驱动体内细胞正常运作所需的大部分ATP,并且在包括心脏和肝脏在内的高能量需求的器官中发挥着重要作用。线粒体质量控制机制,包括线粒体自噬(当线粒体在遭受损伤或应激后出现缺陷时,通常就会发生),有助于确保线粒体网络正常运行。尽管已确定了线粒体自噬的作用,但是到目前为止,关于参与线粒体自噬的不同蛋白的信息仍然是有限的。
为了更深入地了解参与线粒体自噬的组分,这些研究人员使用了多个报告系统和促线粒体自噬触发因子来开展CRISPR/Cas9全基因组筛选。令他们吃惊的是,他们发现ANT是许多细胞类型中线粒体质量控制途径所必需的。在证实他们的发现的后续测试中,他们发现ANT促进了线粒体自噬,但这与它在产生ATP中的作用无关,这表明这种蛋白具有两种不同的功能。
基于这一发现,这些研究人员设计出研究ANT基因突变是否影响线粒体自噬的测试方法。他们发现,ANT中的致病突变清除了它与将蛋白从线粒体内膜重定位到线粒体基质中所必需的蛋白复合物结合的能力,从而最终抑制了线粒体自噬。进一步的研究表明通过遗传手段剔除ANT可以抑制线粒体自噬。他们还发现,缺乏ANT的小鼠具有较弱的线粒体自噬能力,因而经历了非典型线粒体的大量积累。
Arany说,“我们非常吃惊地发现ANT参与线粒体自噬,这揭示出它具有两种独立的功能---ATP产生和线粒体质量控制。尽管目前没有改善线粒体质量控制的灵丹妙药,但是我们希望我们的发现为今后的研究工作打下基础,从而有助于研究如何抵消对线粒体自噬的抑制,并且最终阻止这些突变引起疾病。”
8. Nature:揭示一种阻止蛋白聚集物在线粒体中聚集的新机制
doi:10.1038/s41586-019-1307-z.
蛋白聚集物对线粒体功能是有害的,因而会破坏向它们的宿主细胞提供化学能。在一项新的研究中,来自德国慕尼黑大学等研究机构的研究人员描述了一种阻止这些蛋白聚集物在线粒体中聚集的蛋白复合物。相关研究结果近期发表在Nature期刊上,论文标题为“Structure and function of Vms1 and Arb1 in RQC and mitochondrial proteome homeostasis”。
作为一种细胞器,线粒体为高等生物中的细胞提供它们的代谢和功能维持所必需的化学能。此外,许多必需代谢物的生物合成发生在这些细胞器中。因此,必须快速检测和修复它们的功能出现的任何扰动---比如,错误折叠蛋白的异常堆积。
在慕尼黑大学生物医学中心的Walter Neupert、慕尼黑大学基因中心的Roland Beckmann和日本仙台大学的Toshifumi Inada的领导下,这些研究人员如今阐明了细胞抑制前往线粒体的毒性蛋白聚集物形成的机制之一。如果不加以抑制的话,这些蛋白聚集物就会切断为不可或缺的细胞功能提供的能量。
蛋白由称为核糖体的细胞器合成。这些RNA-蛋白复合物(即核糖体)对从细胞核输出的确定着特定蛋白氨基酸序列的信使RNA(mRNA)蓝图进行解码。核糖体从确定的起始位点开始依次附着于mRNA分子上。这允许每个mRNA分子以装配线方式编程每种特定蛋白的许多拷贝的合成。如果这个过程受到中断而且前面的核糖体发生停滞,那么后面的核糖体堆积在后面。在这些条件下,它们附着的不完整蛋白(译者注:指的是因核糖体停滞,核糖体上的蛋白未完整合成)能够容易地彼此相互作用以形成蛋白聚集物。为解决此类交通拥堵,细胞激活“核糖体相关质量控制(ribosome-associated quality control, RQC)机制”。RQC途径的作用机制是让停滞的核糖体的两个亚基(60S大亚基和40S小亚基)分离开来(从而释放mRNA)并且将丙氨酸(A)和苏氨酸(T)附着到60S亚基上不完整蛋白的停滞羧基末端(译者注:指的是因核糖体停滞,核糖体在mRNA的特定局部大量堆积,核糖体上的新生蛋白未能完整合成,这就意味着在发生核糖体停滞后,新生蛋白的羧基末端停滞了,也就是羧基末端不再添加新的氨基酸)上。这样,这种不完整的蛋白从核糖体中释放出来,“CAT尾巴”标志着它随后会被降解。
然而,就线粒体蛋白而言,情况就更加复杂了。大多数受到抑制或发生停滞的线粒体蛋白是由附着在线粒体膜孔上的核糖体在细胞质中形成的,这些新生的线粒体蛋白通过这些孔直接进入这种细胞器。这种合成与线粒体摄取的紧密连接阻碍了CAT标记的蛋白在细胞质中的释放和裂解。此外,CAT标记的蛋白本身具有增加的聚集倾向,因此它们的输入对线粒体功能尤其有害。
幸运的是,细胞已进化出一种专门用于线粒体蛋白的RQC替代形式。这项新研究的作者之前已证实Vms1蛋白在这一途径中发挥着关键作用,但这种RQC模式的确切机制尚不清楚。Beckmann说,“通过将基于低温电镜的结构分析与生化、分子生物学和遗传学实验相结合,我们如今成功地阐明了Vms1的工作机制。”Vms1能够解离核糖体并释放出停滞的蛋白,不论这种停滞的蛋白是否携带CAT序列。因此,它有效地抵消了添加到线粒体蛋白上的CAT序列,从而降低了蛋白聚集的风险以及对线粒体和细胞功能的不利影响。此外,Beckmann及其同事们发现了一种之前未被描述的在这整个过程中发挥作用的蛋白:Arb1,并展示了它与Vms1之间如何相互作用。这些新的研究结果可能有助于更好地了解各种疾病,包括代谢疾病和神经退行性疾病,这些疾病与线粒体功能的损害有关。
9. Nature:线粒体裂殖需要蛋白DRP1,但不需要动力蛋白
doi:10.1038/s41586-019-1296-y.
线粒体裂变(mitochondrial fission,有时也译作线粒体分裂)是维持线粒体网络所必需的,并且依赖于一种称为动力蛋白相关蛋白1(dynamin-relatedprotein 1, DRP1,也称为DNM1L)的GTP酶。DRP1形成螺旋寡聚体,包裹线粒体外膜并将其分裂。最近,有人提出DRP1不足以进行线粒体裂变,另一种称为动力蛋白-2(dynamin-2, DNM2, 也称为DYN2)的GTP酶是线粒体分裂复合物的重要组成部分。
在一项新的研究中,来自德国哥廷根大学医学中心的研究人员报道缺乏所有三种哺乳动物动力蛋白(DNM1、DNM2和DNM3)的小鼠成纤维细胞(下称动力蛋白三重敲除细胞)以及仅敲降DNM2的细胞既没有表现出线粒体裂变或过度融合缺陷,但是敲降DRP1可很容易检测到这些缺陷,即便在动力蛋白三重敲除细胞中,也是如此。相关研究结果发表在2019年6月20日的Nature期刊上,论文标题为“Mitochondrial fission requires DRP1 but not dynamins”。
鉴于线粒体和过氧化物酶体具有相同的裂变机制,这些研究人员研究了过氧化物酶体裂变,结果观察到与线粒体裂变相同的情形。
因此,这些研究结果表明DRP1对于线粒体和过氧化物酶体裂变是必需的,而哺乳动物动力蛋白DNM1、DNM2和DNM3则并不是必需的。(生物谷Bioon.com)
版权声明 本网站所有注明“来源:生物谷”或“来源:bioon”的文字、图片和音视频资料,版权均属于生物谷网站所有。非经授权,任何媒体、网站或个人不得转载,否则将追究法律责任。取得书面授权转载时,须注明“来源:生物谷”。其它来源的文章系转载文章,本网所有转载文章系出于传递更多信息之目的,转载内容不代表本站立场。不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
87%用户都在用生物谷APP 随时阅读、评论、分享交流 请扫描二维码下载->
